Большинство слизистых оболочек, особенно слизистые оболочки I типа в кишечнике и легких, по своей природе являются «хрупкими», так что они легко могут быть повреждены продуктами патогенов во время развития воспалительных реакций. По этой причине иммунитет слизистой оболочки в значительной степени формируют секреторные антитела и, в первую очередь, секреторный иммуноглобулин класса А (sIgA). Эти антитела являются ключевыми компонентами слизи и других жидкостей организма барьерных тканей, таких как слюна и слезная жидкость, и защищают от патогенных микроорганизмов. Хотя sIgM присутствует в некоторых жидкостях организма, подавляющее большинство секреторных антител – это sIgA. Мыши, неспособные продуцировать sIgA из-за нарушения С альфа-экзона или дефектов синтеза J-цепи, демонстрируют повышенную восприимчивость к инфекции и неблагоприятным изменениям состава микробиоты. Секреторный IgA имеет несколько особенностей, которые делают его идеальным для защиты слизистой оболочки. Во-первых, sIgA конститутивно локализуются в слизи, для нейтрализации практически любого патогена или токсина, способного вступить в контакт с эпителием. Во-вторых, независимо от антигенной специфичности, углеводные фрагменты С-доменов молекул sIgA могут связываться с молекулами адгезии, экспрессируемыми многими патогенами. В-третьих, по крайней мере, в кишечнике около половины всех антител sIgA обладают способностью перекрестно реагировать с большим количеством эпитопов. Это означает, что более широкому спектру угроз можно противостоять с помощью меньшего количества антител. В-четвертых, sIgA не являются эффективными активаторами комплемента, что снижает вероятность активации этого каскада и инициации повреждающего воспаления при формировании комплексов антиген-антитело. В-пятых, sIgA обладает высокой устойчивостью к широкому спектру протеаз хозяина и микробов, в том числе, в кишечнике млекопитающих.
Механизмы выработки sIgA слизистой оболочки
Энтероциты, взаимодействуя с комменсальными бактериями, продуцируют цитокины, которые индуцируют продукцию В клетками sIgA
Выработка sIgA в организме намного превышает выработку антител любого другого изотипа. В среднем, в кишечнике взрослого человека ежедневно синтезируется около 2-3 граммов sIgA. sIgA-продуцирующие плазматические клетки присутствуют в кишке даже в отсутствие патогенов из-за влияния микробиоты на В-клетки слизистой. Постоянная продукция sIgA держит численность комменсалов под контролем, защищая слизистые оболочки и поддерживая гомеостаз.
В случае Т-зависимых реакций, cDC способствуют формированию высокоаффинных IgA не только к комменсальной микробиоте, но и к патогенам и их токсинам. cDC в Пейеровых бляшках регулируют процессы переключения класса синтезируемых антител на IgA, а также экспрессию рецепторов «хоуминга» в ткани слизистых на IgA продуцирующих плазматических клетках и их предшественниках.
Энтероциты слизистых оболочек также играют важную роль в продукции sIgA (Рис.94). Когда энтероциты с помощью PRR (особенно TLR2) связываются с комменсальными бактериями, это вызывает продукцию цитокинов BAFF (от англ. «B-cell-activating factor belonging to the TNF family»), APRIL (от англ. «a proliferation inducing ligand») и IL-10, которые индуцируют в 80% активированных В-клеток lamina propria переключение изотипа иммуноглобулинов именно на IgA.
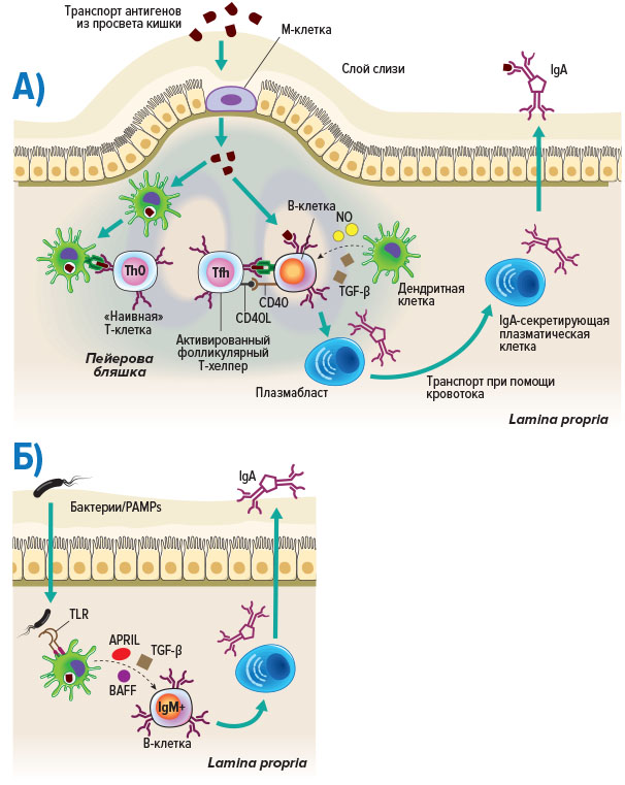
A. Переключение на IgA происходит при связывании APRIL (DCs и эпителий) с TACI –рецептором на В-клетках. Эпителиальные клетки кишечника также продуцируют TSLP, который стимулирует продукцию APRIL дендритными клетками.
Эпителий и DCs продуцируют TGF-β. TLR лиганды, производимые комменсальными бактериями также индуцируют выработку NO, который тоже способствует выработке IgA. Наконец, выработка IgA зависит от ретиноевой кислоты, которая производится клетками эпителия и DCs, экспрессирующими ретинолдегидрогеназу RALDH.
B. Переключению класса IgA способствуют сигналы от TLR.
Энтероциты также выделяют TSLP, который усиливает выработку BAFF, APRIL и IL-10. Кроме того, cDC2, которые находятся в куполе Пейеровых бляшек мышей, секретируют высокие уровни IL-10. Презентация DCs антигенов в присутствии IL-10 индуцирует поляризацию «наивных» Т лимфоцитов в Tfh; регуляторные Т-клетки, секретирующие TGFβ и IL-10; и Th2-эффекторы, синтезирующие IL-4, IL-5 и IL-10. Одновременно поляризация в Th1 и продукция IFNg подавляются. При распознавании TLR5 флагеллина, DCs начинают экспрессировать фермент ретинолдегидрогеназу, который конвертирует витамин А в ретиноевую кислоту. Ретиноевая кислота, TGFβ и IL-10 направляют переключение изотипов иммуноглобулинов в активированных В-клетках в сторону IgA. В-лимфоциты циркулируют через лимфу и кровь в эффекторные сайты слизистой оболочки (напр. мезентериальные лимфоузлы). В этих местах активированные В-клетки завершают свою дифференцировку в зрелые IgA-продуцирующие плазматические клетки.
Плазматические клетки секретируют полимерный IgA вблизи базолатеральной поверхности эпителиальных клеток, выстилающих кишечник, а также клеток, образующих экзокринные железы. Полимерный IgA связывается с поли-иммуноглобулиновым рецептром или pIgR (от англ. «poly-Ig receptor») на базальной поверхности клеток эпителия в эффекторных участках слизистой оболочки и переносится при помощи трансцитоза на апикальную поверхность клеток. При экзоцитозе pIgR расщепляется таким образом, что секреторный компонент остается прикрепленным к антителу, что приводит к высвобождению комплекса sIgA в слизь (Рис.95)
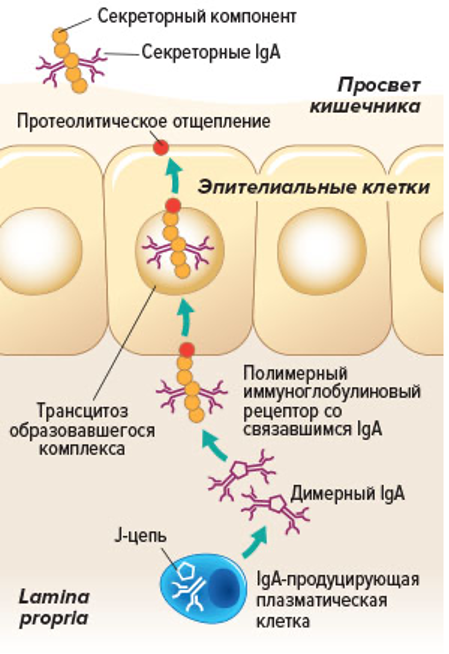
IgA транспортируются с базальной на апикальную поверхность эпителия. IgA продуцируется в ламина проприа в форме димера, Fc-фрагменты мономеров, которого ковалентно связаны с J цепью. Димерный IgA транспортируется из ламина проприа в просвет кишечника. Этот транспорт осуществляется с помощью poly-Ig рецептора на базальной и апикальной поверхности эпителиальных клеток. Секреторный компонент связывается с бактериями и снижает их подвижность.
Плазматические клетки мыши продуцируют только один подкласс IgA, но у человека продуцируется IgA1 и IgA2, которые отличаются IgA транспортируются с базальной на апикальную поверхность эпителия. IgA продуцируется в ламина проприа в форме димера, Fc-фрагменты мономеров, которого ковалентно связаны с J цепью. Димерный IgA транспортируется из ламина проприа в просвет кишечника. Этот транспорт осуществляется с помощью poly-Ig рецептора на базальной и апикальной поверхности эпителиальных клеток. Секреторный компонент связывается с бактериями и снижает их подвижность.
Плазматические клетки мыши продуцируют только один подкласс IgA, но у человека продуцируется IgA1 и IgA2, которые отличаются некоторыми своими свойствами. В то время как выработка IgA1 происходит как системно, так и на слизистых, IgA2 присутствует главным образом на слизистых поверхностях (особенно в дистальном отделе кишечника и урогенитальном тракте). Благодаря более короткой области шарнира IgA2 более устойчив, чем IgA1, к деградации протеазами, что позволяет этому антителу обеспечивать устойчивую защиту в суровой биохимической среде.
Th17 тип адаптивного ответа обеспечивает защиту от внеклеточных патогенов, например, грибов Candida albicans или бактерий Salmonella typhimurium, C.rodentium и Yersinia enterocolitica. Поддержание Th17 ответа в кишечнике зависит от сигналов микробиоты, в частности, сегментированных нитевидных бактерий. В исследованиях in vitro было показано, что активированные зрелые DCs GALT направляют поляризацию «наивных» активированных CD4+ Т-клеток преимущественно в сторону Th17. Недавние исследования на мышах показали, что микробиота может оказать влияние на формирование иммунитета в GALT. Например, мыши, выращенные в стерильных условиях, не имеют микробиоты в кишечнике и обладают неразвитой GALT, аномально высоким количество Th2-клеток и аномально низкими уровнями Th1- и Th17-клеток. На энтероцитах у этих животных наблюдается снижение экспрессии TLR, и их DCs не могут формировать трансэпителиальные отростки.