Интерес к регуляторным Т-лимфоцитам как основной популяции клеток периферической крови, способной подавлять развитие воспалительных реакций по всем трем описанным выше типам, не угасает уже многие годы. Снижение же функциональной активности Treg приводило к нарушению толерантности к собственным антигенам организма и развитию аутоиммунных патологических состояний. Популяция Treg не является однородной. Описаны как минимум две субпопуляции, циркулирующие в периферической крови и принципиально различающиеся по происхождению. «Тимические» или «натуральные» регуляторные Т-лимфоциты (от англ. «thymus derived» или «natural» Treg, nTregs или tTregs) проходят дифференцировку в тимусе в ходе антиген-независимой стадии созревания Т-клеток. nTregs обладают фенотипом «наивных» клеток периферической крови — CD45RA+CD45R0–CD62L+CCR7+.
Другая популяция «периферические» или «индуцибельные» или «адаптивные» Treg (от англ. «peripherally derived», «induced» или «adaptive» Treg, iTregs) – регуляторные Т-клетки формируется в процессе антиген-зависимой дифференцировки в периферических лимфоидных органах. В 2013 году была предпринята попытка стандартизировать номенклатуру Treg, и было предложено Treg, дифференцирующиеся в тимусе, обозначать как nTreg, а Treg, формирующиеся из Th0 на периферии, в лимфатических узлах, как pTreg. Название iTreg (от англ. «in vitro–induced» Treg) было предложено употреблять для обозначения регуляторных Т-клеток, которые были получены в ходе дифференцировки в условиях in vitro, чтобы отличать их от pTreg, формирующихся in vivo. Эффекторные молекулы регуляторных Т-лимфоцитов представлены на рис.84.
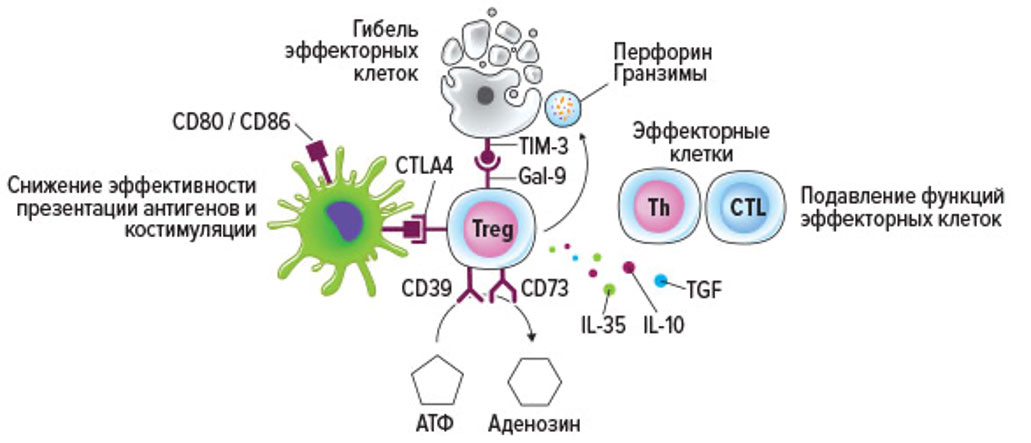
Мишенями Treg являются все клетки врожденного (тканевые макрофаги, антиген-презентирующие клетки, натуральные киллеры) и приобретенного иммунитета (эффекторные цитотоксические Т-лимфоциты и Т-хелперы, а также В-лимфоциты). Для реализации функций Treg используют различные механизмы, которые традиционно разделяют на «бесконтактные» (опосредованные действием высвобождаемых регуляторными Т-клетками различных растворимых молекул, диффундирующих в тканевые жидкости) и «контактные» (опосредованные взаимодействием рецепторов Treg и поверхностных структур на клетке-мишени).
«Бесконтактные» механизмы супрессии Treg.
Ключевыми иммунорегуляторными гуморальными факторами Treg человека являются IL-10, TGFb и IL-35, а также внеклеточный аденозин. Аденозин образуется путем гидролиза внеклеточного АТФ ферментами CD39 и CD73, локализованными на цитоплазматической мембране Treg.
IL-10 является белком с молекулярной массой около 18 кДа, который, помимо Treg, синтезируют моноциты, дендритные клетки, нейтрофилы, различные популяции Th (например, Th17 и Th22, как уже отмечалось ранее) и некоторые популяции В-лимфоцитов (значение этих популяций В-клеток будет рассмотрено в следующей главе данной монографии). Ключевыми функциями данного цитокина являются:
– угнетение продукции провоспалительных цитокинов и, в первую очередь, IL-12, играющего ведущую роль в «поляризации» Th0 в сторону Th1. Это приводит к снижению продукции IFNγ и формирования эффекторных Th1;
— за счет блока эффектов IFNγ, усиление дифференцировки тканевых макрофагов в М2, которые также являются продуцентами противовоспалительных цитокинов – IL-10 и TGFb;
– увеличение эффективности фагоцитоза и элиминации клеточного дебриса в очаге воспаления,
– снижение эффективности взаимодействия между АПК и антиген-распознающими клетками, путем блокировки передачи сигнала от TcR в ядро лимфоцитов. Снижение экспрессии ко-стимулирующих молекул CD80 и CD86 и комплексов антиген-MHC II, что существенно ограничивает пролиферативную активность, созревание и дифференцировку клона антиген-специфических лимфоцитов (как Т-хелперов, так и цитотоксических Т-лимфоцитов);
TGFb является белком с молекулярной массой около 25 кДа, который секретируется в виде неактивного пептида, состоящего из собственно TGFb и пептида LAP (от англ. «Latency-Associated Peptide»). TGFb снижает пролиферативную активность активированных Т-лимфоцитов за счет подавления синтеза и секреции IL-2 и подавляет их дифференцировку в эффекторные клетки (как Т-хелперов, так и эффекторных цитотоксических Т-лимфоцитов). TGFb, стимулируя экспрессию транскрипционного фактора FOXP3, способствует преимущественной поляризации регуляторных Т-лимфоцитов. Под действием TGFb снижается уровень активации тканевых макрофагов, дендритных клеток и натуральных киллеров, равно как и пролиферация активированных В-лимфоцитов.
IL-35 принадлежит к семейству IL-12 и является гетеродимерным белком, состоящим из двух субъединиц – p35 и Ebi3. В отличие от остальных членов данного семейства (IL-12, IL-23 и IL-27), обладающих провоспалительными свойствами, IL-35 обладает выраженной иммуносупрессорной активностью. Основными функциями данного цитокина являются подавление пролиферативной активности «наивных» Т-лимфоцитов и «поляризация» Th0 в сторону Treg, продуцирующих большое количество IL-35 и получивших еще название «iTr35». Считается, что именно IL-35 является мощнейшим фактором, ограничивающим провоспалительную функциональную активность Th17, так как он способен подавлять дифференцировку Th в сторону эффекторных Th17.
Аденозин подавляет адгезию лейкоцитов к эндотелиальным клеткам микроциркуляторного русла, снижает продукцию супероксид анионов нейтрофилами и снижает синтез и секрецию провоспалительных цитокинов клетками различного происхождения. Аденозин способствует продукции противовоспалительного IL-10 моноцитами, а также запускает продукцию VEGF – мощного индуктора ангиогенеза и сосудистой проницаемости. Большинство из этих эффектов аденозина реализуются через различных представителей семейства A2 рецепторов. Аденозин подавляет активность эффекторных Т-лимфоцитов, ингибирует дифференцировку Th1 и Th2 за счет снижения уровня пролиферативной активности и продукции IL-2, что выражается еще и в снижении продукции эффекторных цитокинов, свойственных Th1 и Th2 линиям дифференцировки Т-хелперов. Он также угнетает активность (проявление цитолитических свойств или синтез цитокинов), и других популяций Т-клеток.
Под действием аденозина снижается экспрессия ко-стимулирующих молекул – CD2 и CD28 – активированными Т-лимфоцитами. Для самих Treg аденозин является мощнейшим аутокринным фактором активации. Под его воиянием на поверхности Treg увеличивается плотность супрессорных молекул CTLA-4, CD39 и CD73, а также уровень экспрессии CD25. Также было отмечено ,что после культивирования in vitro в присутствии аденозина происходит повышение доли Treg, что указывает на активацию процессов пролиферации данной популяции клеток. «Контактные» механизмы иммуносупрессии основаны на взаимодействии рецепторов Treg с их лигандами на клетке-мишени.
CTLA-4 (от англ. «Cytotoxic T-lymphocyte–associated antigen 4», или CD152) экспрессируется Treg. Известно, что CTLA-4 имеет высокую плотность экспрессии на Treg и ингибирует иммунный ответ. Главными лигандами CTLA-4 являются ко-стимулирующие молекулы CD80 и CD86. Эти молекулы также являются лигандами активационной костимулирующей молекулы CD28. Но аффинность связывания CTLA-4 с лигандами, значительно выше, чем таковая для CD28. Именно поэтому основной функцией CTLA-4 является «экранирование» CD80 и CD86 от активированных Т-клеток, экспрессирующих CD28.
В результате взаимодействия CTLA-4 с лигандами происходит интернализация CD80 и CD86 антиген-презентирующей клеткой, что существенно снижает эффективность презентации антигена и угнетает формирование клона антиген-специфических Т-клеток. С другой стороны, Treg через взаимодействие CTLA-4 с CD80/86 регулируют созревание дендритных клеток.
LAG3 (от англ. «Lymphocyte-Activation Gene 3» или CD223). Основным лигандом LAG3 являются молекулы МНС II класса на АПК, причем аффиность взаимодействия LAG3-MHC II существенно выше такового комплекса CD4-MHC II, что способствует снижению активации Т-хелперов и блокирует развитие воспаления по всем трем типам.
Treg способны вызывать апоптоз эффекторных клеток при помощи двух основных механизмов. Во-первых, Treg индуцируют апоптоз за счет секреции в окружающее пространство перфорина и гранзимов, как это показано для натуральных киллеров и эффекторных цитотоксических Т-лимфоцитов. Во-вторых, индукции апоптоза клеток мишеней реализуется за счет экспрессии на поверхности Treg галектина-9 (Gal-9) – β-галактозидаза-связывающего белка, способного связываться с TIM-3 (от англ. «T cell immunoglobulin and mucin domain 3» или CD366) на поверхности зрелых эффекторных клеток. Результатом такого взаимодействия является активации каспаз в клетке-мишени. Кроме того, за счет Gal-9 осуществляется подавление пролиферации антиген-специфических Т-клеток.
В настоящее время существует множество различных определений понятия «иммунологическая толерантность», некоторые из которым приводятся ниже. Иммунологическая толерантность:
— отсутствие иммунного ответа при наличии в организме антигенов (толерогенов), доступных лимфоцитам;
— специфическая ареактивность к какому-либо антигену, индуцированная предшествующим контактом с этим антигеном (на уровне организма или иммунной системы);
— это отсутствие активации лимфоцитов (и, соответственно, продукции эффекторных молекул) при наличии специфического антигена (на уровне отдельных клонов клеток иммунной системы).
В чем состоит биологический смысл иммунологической толерантности?
- В ходе развития лимфоцитов в центральных органах иммунной системы, в результате реаранжировки генов возникают уникальные Т- и В-клеточные рецепторы. Поэтому каждая клетка – Т- или В-лимфоцит – УНИКАЛЬНА, что является ключевым отличием этих клеток от клеток врожденного иммунитета.
- Так как реаранжировка генов рецепторов носит «случайный» характер, образующие при этом Т- и В- лимфоциты способны к распознаванию любых антигенов, включая антигены собственного организма – то есть «аутоантигены». Именно это может приводить к развитию аутоиммунных заболеваний. Если по каким-то причинам, АПК презентирует аутоантигены при наличии 2 и 3 сигналов (костимуляции и цитокинового) «свое» распознается системой приобретенного иммунитета как «чужое».
- Именно поэтому при созревании Т- и В-клеток необходимы механизмы «селекции» — индукция «центральной» толерантности, формирование «периферической» толерантности или контроль за потенциальными «беглецами», В-клетками, которые изменяют специфичность своего рецептора в ходе развития иммунного ответа.
Центральная (иногда называет «естественная») толерантность — это совокупность механизмов, направленных на уничтожение аутореактивных клеток в центральных лимфоидных органах (тимус и красный костным мозг) в ходе их (клеток) созревания и дифференцировки. ОДНАКО в ходе этого процесса часть аутореактивных клеток может избегать апоптоза. К механизмам индукции центральной толерантности относятся — негативная селекция клеток; редактирование рецепторов (для В-лимфоцитов), а также формирование «натуральных» или «тимических» Treg, которые способны распознавать собственные антигены и блокировать развитие иммунного ответа на них при помощи механизмов, которые детально описаны нами в разделе, посвященном данной популяции Т-лимфоцитов.
Центральная иммунологическая толерантность обусловлена отрицательной селекцией аутореактивных клонов лимфоцитов. и развивается в процессе онтогенеза за счет уничтожения аутореактивных клонов Т- и В-лимфоцитов. Классическим примером формирования толерантности к собственным антигенам организма является процесс «негативной» селекции тимоцитов (будущих Т-лимфоцитов) в тимусе. Так, в ходе позитивной селекции сигнал к выживанию получают клетки, способные распознавать молекулы МНС в ассоциации с собственными антигенами организма и способные, как следствие, проводить сигнал от TсR. Остальные тимоциты не получают необходимый сигнал от TcR и в дальнейшем гибнут путем апоптоза. Клетки, прошедшие положительную селекцию, т.е. сформировавшие функционально-активный Т-клеточный рецептор, проходят следующий этап отбора, получившего название «негативной» селекции.
В ходе этого процесса происходит оценка интенсивности сигнала от TcR, который формируется при распознавании аутоантигенов, то есть собственных антигенов организма, экспрессирующихся на строме тимуса. Ключевая роль в негативной селекции отводится антиген-презентирующим и эпителиальным клеткам. Медуллярные тимические эпителиальные клетки экспрессируют ядерный белок, называемый AIRE (аутоиммунный регулятор, от англ. «autoimmune regulator»), который, как считается, способен на низком уровне индуцировать в тимусе экспрессию множества генов. Белковые продукты этих генов обычно экспрессируются только в определенных периферических органах и тканях, и называются «тканеспецифичные» антигены. Экспрессия в тимусе «тканеспецифичных» антигенов делает их доступными для презентации и распознавания незрелыми Т-клетками.
Возможно, что именно этот механизм обеспечивает негативную селекцию и удаление аутореактивных клонов и формирования пула тимических регуляторных Т-клеток, способных подавлять ответ на собственные антигены организма в периферических лимфоидных органах. Косвенным подтверждением такого механизма селекции и роли AIRE в формировании центральной толерантности является тот факт, что мутации в гене, кодирующем белок AIRE, приводят к развитию аутоиммунного полиэндокринного синдрома.
В ходе негативной селекции высокоаффинное связывание TcR с комплексом аутоантиген-МНС генерирует сильный сигнал, запускающий в тимоцитах процесс апоптоза. Слабый сигнал свидетельствует о низкой аффинности связывания TcR с аутоантигеном, что дает возможность клетке продолжить свое развитие. В то же время «промежуточные» значения этого сигнала способствуют формированию тимических или натуральных регуляторных Т-лимфоцитов. Следует особо подчеркнуть, что тонкие механизмы негативной селекции в настоящее время недостаточно хорошо изучены, и наши представления об этом носят в значительной степени теоретический характер.
Периферическая толерантность — это совокупность механизмов, направленных на уничтожение аутореактивных клеток в периферических лимфоидных органах (лимфатические узлы, Пейеровы бляшки и т.д.) при инициации иммунного ответа.
Периферическая толерантность — элиминация «беглецов» (Т- и В-клеток) и вновь образованных после соматических гипермутаций аутореактивных В-клеток. Ведущую роль в регуляции механизмов поддержания периферической толерантности играют «ауто-реактивные» Treg. При генетических дефектах, затрагивающих развитие Treg-клеток аутоиммунные реакции проявляются наиболее сильно. Это наблюдается у больных с IPEX-синдромом (врожденные иммунодефицит, X-ассоциированная иммунная дисрегуляция, полиэндокринопатия, энтеропатия), который развивается в результате мутации гена FOXP3/Foxp3 и связан с множественными аутоиммунными реакциями. Как уже нами отмечалось ранее, Foxp3 является ключевым маркером регуляторных Т-клеток, который регулирует формирование в тимусе и поддержание пула Treg на периферии, а также крайне важен для реализации супрессорных функций этих клеток.
Периферическая толерантность реализуется за счет следующих механизмов:
— делеция клона. Лимфоциты, связавшие антиген вместо активации, погибают в результате апоптоза;
— анергия клона. Лимфоциты, связавшие антиген вместо активации, входят в состояние «неотвечаемости» из-за отсутствия 2 сигнала (ко-стимулирующих молекул) со стороны АПК;
— проявление эффекторных свойств Treg (продукция противовоспалительных цитокинов, образование аденозина и широкий спектр контактных взаимодействий с клетками-мишенями).