Любой повреждающий агент-фактор, который по силе и длительности превосходит адаптационные возможности ткани, может вызвать ответную защитную реакцию организма. Эти факторы принято делить на внешние (экзогенные – микроорганизмы, химические вещества, физические воздействия) и внутренние (эндогенные – появляющиеся в самом организме в результате патологического процесса). Защитная реакция направлена на то, чтобы не допустить распространения патогена, ограничив и уничтожив его на месте проникновения, привлекая факторы всего организма (в отличии от мукозального иммунитета, где реакции обычно ограничиваются «местными» клетками и молекулами). Это достигается, прежде всего, воспалением.
Воспаление представляет собой скоординированный каскад системных иммунных, эндокринных и неврологических реакций, возникающий, в случае, когда повреждающий агент по силе и длительности превосходит барьерные возможности ткани, проявляется в локальных и системных реакциях, направленных на устранение патогена и максимального восстановления зоны повреждения.
Несмотря на многообразие факторов, вызывающих воспалительную реакцию, закономерности ответа на повреждение, которые происходят в тканях, однотипны.
Воспаление начинает формироваться с процесса распознавания чужеродных веществ. Чужеродным веществом выступают патогенные микроорганизмы, которые имеют молекулярные структуры, отсутствующие в организме человека. PAMP (Pathogen-associated molecular patterns (образы патогенности) чужеродные структуры патогенов). Это пептидогликаны, ЛПС (липополисахариды), липопротеины, липотейхоевая и тейхоевая кислоты, флагеллин, нуклеиновые кислоты вирусов и бактерий, вирусные белки и др. являющиеся составными частями микроорганизма.
Помимо патогенов иммунная система распознает и нейтрализует молекулярные структуры, связанные с повреждением собственных тканей — DAMP (danger-associated molecular patterns (образы опасности), эндогенные вещества клеток (белки теплового шока, белки S100, фибриллы амилоида-ß, дефенсины, кателицидины, галектины, аннексины, тимозины, цитокины (ИЛ-1α и ИЛ-33), хроматин-связанный белок HMGB1 , мочевая кислота и др.), образуемые при повреждении клеток и клеточном стрессе).
Помимо PAMP и DAMP, которые могут распознаваться разными рецепторами, иммунная система способна распознавать и индивидуально формировать рецепторы (иммуноглобулиновые и/или специальные Т-клеточные рецепторы совместно с молекулами MHC II-класса) к генетически чужеродным молекулярным структурам – антигенам (АГ). Это длительный сложный процесс, зависящий от многих факторов, но способный обеспечить распознавание молекулярной структуры любой природы. Принято выделять экзогенные и эндогенные АГ. Экзогенные АГ инфекционные и паразитарные молекулярные структуры вирусов, риккетсий, бактерий, грибов, одно- и многоклеточных паразитов и неинфекционные чужеродные белки; белоксодержащие соединения; антигены и гаптены в составе пыли, пищевых продуктов, пыльцы растений, ряда лекарственных средств перерабатываемые антигенпредставляющими клетками с презентацией АГ. Подобные реакции могут запускать и собственные структуры организма, распознанные как чужеродные (повреждённые белки и содержащие белок собственные клетки, неклеточные структуры и жидкости организма, при конъюгации с ними гаптенов, в результате мутаций, приводящих к синтезу аномальных белков, при сбоях иммунной системы).
В процессе разрушения клеток выходит большое количество DAMP, которые совместно с PAMP распознаются макрофагами, эпителиальными клетками и кератоцитами с формированием в них инфламмасом (цитозольный мультибелковый олигомер ответственный за активацию воспалительных реакций) с активным синтезом провоспалительных цитокинов и инициации пироптоза.
Врожденный иммунный ответ основан на распознавании чужеродных агентов с помощью специальных патоген-распознающих рецепторов (PRR). По функции все PRR могут быть разделены на сигнальные и эндоцитозные.

Первая группа рецепторов позволяет выявлять патоген-ассоциированные молекулярные структуры – PAMP, отсутствующие в организме человека (пептидогликаны, липополисахариды, липопротеины, липотейхоевая и тейхоевая, кислоты, флагеллин, нуклеиновые кислоты вирусов и бактерий, вирусные белки и др.), и/или молекулярные структуры, связанные с повреждением – DAMP – эндогенные вещества клеток, образуемые при гибели клеток и клеточном стрессе (белки теплового шока, белки S100, фибриллы амилоида-ß, дефенсины, кателицидины, галектины, аннексины, тимозины, цитокины ИЛ-1α и ИЛ-33, хроматин-связанный белок HMGB1, мочевая кислота и др.).
Вторая группа рецепторов — эндоцитозные PRR обеспечивает процессы фагоцитоза и последующую доставку патогена в лизосомы (начало адаптивного иммунного ответа) и активирует эффероцитоз (узнавание и поглощение апоптотических клеток).
Помимо этого, выделяют рецепторы врожденной иммунной системы и рецепторы адаптивного иммунитета. Рецепторы врожденного иммунитета одинаковы у разных людей, ограничены по количеству и разнообразию, напротив рецепторы адаптивного иммунитета генерируемые при сборки имеют большого количество рецепторов используя случайные комбинации генов.
Рецепторы врожденной иммунной системы
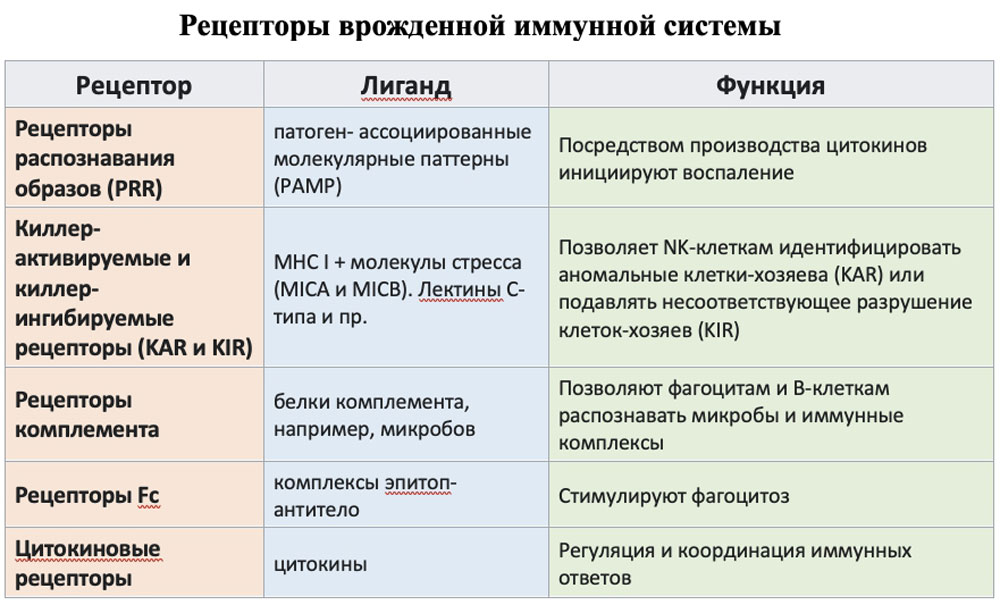
Эпителиальные клетки имеют патоген-распознающие рецепторы, позволяющие выявлять PAMP. Взаимодействие PRR с PAMP и/или DAMP через последовательную активацию адапторных белков, протеинкиназ и транскрипционных факторов, приводит к синтезу и секреции цитокинов (ИЛ-1, -2, -6, -8, -12, ФНО-α, ИФН-γ, ГМ-КСФ). За счет этого происходит активации самих клеток, несущих PRR, значительно усиливается их защитный потенциал (активируется продукция противомикробных пептидов и комплемента, усиливается фагоцитоз, переваривающая активность, продукция активных форм кислорода).
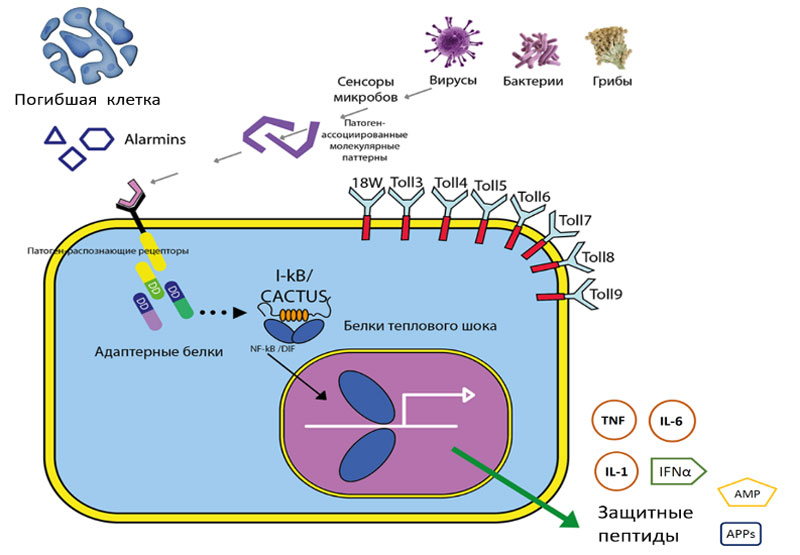
Эпителиальные клетки в основном имеют сигнальные PRR. Однако от их способности синтезировать защитные пептиды, обеспечить доступ лейкоцитов к тканям зависит исход развития заболевания.
Подобные PRR экспрессируются и на других клетках, находящихся в тканях и, прежде всего, на тканевых гранулоцитах (тучные клетки, базофилы, эозинофилы), системе мононуклеарных фагоцитов (моноциты, макрофаги и дендритные клетки), группе «тканевых» лимфоцитов и других неиммунных клеток (эндотелиальные клетки, фибробласты и др.). Именно активный синтез цитокинов привлекает периферические нейтрофилы, эозинофилы, базофилы и тучные клетки к субэпителиальным областям кишечника и ускоряет активацию и дифференцировку местных лимфоцитов.
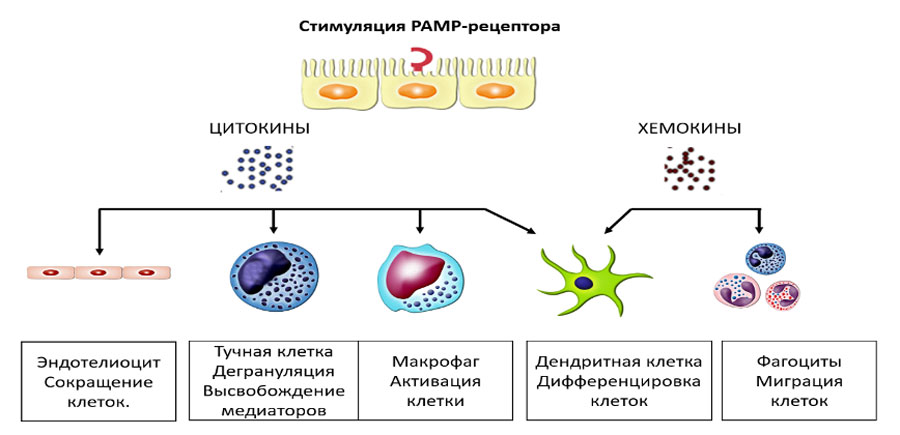
Процесс взаимодействия между патогенными молекулами и иммунной системой происходит по рецепторному типу. В качестве лиганда выступает молекулярные вещества (PAMP, DAMP, АГ), который образует комплекс с клеточным рецептором, отвечающим за обнаружение такого лиганда.
После чего в клетке происходят серия биохимических реакций, инициируемых стимулом (первый мессенджер), передаваемым внутрь клетки через вторичные мессенджеры, которые усиливают сигнал и направляют его эффекторным молекулам, заставляя клетку реагировать на первоначальный стимул в виде биохимических, физиологических или фармакологических эффектов.
Происходит активация большого количества клеток с формированием сосудистой и клеточной реакции. На повреждение отвечают все элементы ткани: микроциркуляторные единицы (артериолы, капилляры, венулы), соединительная ткань, тучные и нервные клетки. Формируется воспаление.
Сосудистая реакция начинается со спазма сосудов – кратковременной реакции, которая переходит в фазу артериальной гиперемии (более длительная). Гиперемия – это усиленное кровенаполнение ткани за счет увеличенного притока крови: возрастает скорость кровотока, повышается давление в сосудах и интенсивность обмена веществ в капиллярах. Отсюда внешние признаки воспаления на этой фазе – покраснение, местный жар (повышение температуры), боль, вызванная действием медиаторов. Уже на этой стадии начинается процесс экссудации.
Под действием медиаторов происходит выход жидкой части плазмы за пределы сосуда — экссудация. Экссудат содержит большое количество белка в связи с нарушением проницаемости сосуда. Он сдавливает венулы и происходит смена артериальной гиперемии на венозную. Чем больше экссудата, тем более выражены явления венозного застоя. Венозная гиперемия постепенно переходит в венозный стаз. Именно в этой фазе происходят значительные изменения поврежденной ткани – так называемые явления вторичного повреждения. Любой венозный застой сопровождается гипоксией: переход на анаэробный процесс окисления – гликолиз, возникновение ацидоза за счет недоокисленных продуктов, т.е. те изменения, которые характерны для первичного повреждения. Накопление кислых продуктов в фазу венозного застоя достигает колоссальных количеств. Наблюдается резко выраженный ацидоз (сдвиг рН до 6,0 – 5,8), а такой сдвиг рН уже непереносим клетками, и они погибают. В центре очага воспаления возникает некроз. При незначительном повышении концентрации водородных ионов (на периферии очага воспаления), нелетальных повреждений клеток незначительный сдвиг рН стимулирует разрастание грануляционной ткани – образуется грануляционный вал на периферии, здоровая ткань отграничивается от поврежденной. Она богата фиксированными макрофагами, способна поглощать поврежденные клетки, токсины, очищая очаг.
Вторичное повреждение также проявляется гиперосмией и гиперонкией. Развитие гиперосмии определяется усиленным катаболизмом и распадом тканей. Распад белковых частиц, жиров, углеводов, выброс калия из клеток с усилением диссоциации солей создают высокую осмотическую концентрацию – гиперосмию. Гиперонкия – увеличение концентрации белков за счет распада ткани, экссудации плазменных белков из сосудов с нарушенной проницаемостью. Эти явления создают порочный круг, усиливая процесс экссудации. Белки как бы притягивают воду, а гиперосмия является повреждающим фактором, который повышает проницаемость стенки сосуда.
При экссудации изменяются биологические свойства крови – увеличивается ее вязкость, кровоток замедляется, усиливаются процессы тромбообразования, наблюдается краевое стояние лейкоцитов, которые выстраиваются вдоль сосудистой стенки, а затем наблюдается их миграция в очаг воспаления. Изменение спектра плазменных белков (выход альбумина, повышение концентрации γ-глобулинов, простагландинов и других медиаторов) влияет на состав мембран, повышает ригидность, преобразует поверхностное натяжение мембран эритроцитов, что усиливает их способность к агрегации (причина ускореного СОЭ).
Тромбоциты тоже приобретают способность к агрегации, но в отличие от эритроцитов этот процесс идет на поверхности сосудистой стенки, в месте ее повреждения. При воспалении происходит ее повреждение, количество простациклина, который предотвращает адгезию и агрегацию тромбоцитов, уменьшается, начинаются процессы адгезии и агглютинации тромбоцитов. Из тромбоцитов выделяются тромбоксаны – мощные стимуляторы процессов адгезии и агрегации. В нормальных условиях простациклин-тромбоксановая система уравновешена. При воспалении происходит активация фактора Хагемана, уменьшение содержания гепарина, что ведет к коагуляции и множественным процессам тромбообразования в очаге воспаления.
Экссудация способствует отграничению очага воспаления, препятствует оттоку токсинов, микробов, распавшихся тканей. В составе экссудата в поврежденную ткань выходят биологические активные вещества, медиаторы, которые способны нейтрализовать токсины, защитные белки, АТ, лейкоциты.
За счет увеличения проницаемости сосудистой стенки происходит проникновение не только белков, но и клеток – клеточная миграция. При этом различные популяции мигрирующих клеток появляются в очаге воспаления в определенном порядке. Первыми, как правило, прибывают нейтрофилы (через 1,5-2 часа). В течение первых суток их популяция преобладает среди пула клеток воспалительного экссудата.
На вторые сутки в очаг начинают поступать мононуклеарные фагоциты и лимфоциты. Позже других мигрируют цитотоксические Т-клетки и В-лимфоциты. Следует отметить, что, как и многие другие процессы при воспалении, миграцию не всегда удается трактовать однозначно. Она изменчива и зависит от целого ряда факторов как эндотелиального, так и лейкоцитарного происхождения. Миграция зависит и от анатомической области, где протекает воспаление, и от присутствия хемотаксических молекул, и от наличия различных цитокинов в тканях (прежде всего хемокинов), и от характера активации мигрирующих клеток (наличие на их поверхности рецепторов) и т.д. В процессах миграции можно выделить несколько основных этапов (рис. 59).
Адгезия лейкоцитов на эндотелии сосудов, осуществляемая за счет молекул межклеточной адгезии (связанные с плазматической мембраной белки, обеспечивающие механическое взаимодействие клеток друг с другом). Выделяют: интегрины — молекулы, функционирующие как клеточно-субстратные, так и межклеточные адгезивные рецепторы; адгезивные рецепторы суперсемейства иммуноглобулинов (ICAM), которые участвуют в межклеточной адгезии, заживлении ран и иммунном ответе; селектины (-L, -Р и -Е) — обеспечивает адгезию лейкоцитов к эндотелиальным клеткам; адрессины – находятся на мембране эндотелиоцитов — лиганды для селектинов, с которыми обеспечивают
адгезию клеток к стенке сосуда, и дальнейшего проникновения в очаг поражения; кадгерины — кальцийзависимые гомофильные межклеточные адгезивные белки; хоминговые рецепторы — молекулы, обеспечивающие попадание лимфоцитов в специфическую лимфоидную ткань.
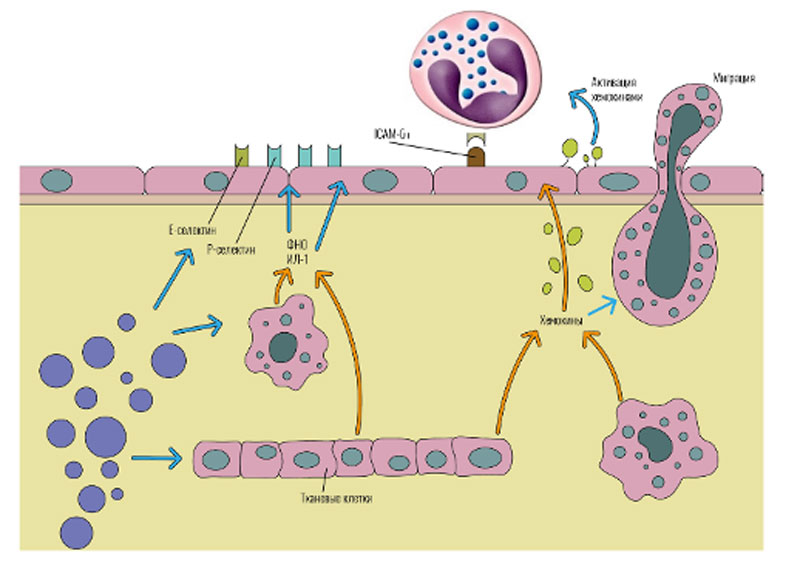
Трансэндотелиальная миграция лейкоцитов, осуществляемая под действием медиаторов воспаления, взаимодействия интегринов и молекул адгезии, клетки преодолевают эндотелиальный слой за счет повышенной сосудистой проницаемости и тока жидкости из сосуда в ткань, а базальные мембраны за счет лизосомальных протеиназ, растворяющие ее, и катионных белков, изменяющие ее коллоидное состояние.
Хемотаксис – направленная миграция в межклеточном пространстве за счет градиента концентрации хемотаксических веществ в очаге воспаления (продукты протеолиза тканей) и разностью потенциалов между отрицательно заряженным лейкоцитом и положительным зарядом ткани.
В процессе развития воспаления клетки участники синтезируют и секретируют большое количество цитокинов, обладающих как про- так и противовоспалительным действием.
Именно от их баланса зависит исход развития патологического процесса как в ранний, так и в отсроченный период (рис 60), и, в конечном итоге, уровень системного воспалительного ответа (СВО).
В последнее десятилетие достигнуты значительные успехи в понимании патофизиологических изменений в организме во время критических состояний. Продолжает формироваться представление о СВО, уточняются и дополняются критерии его оценки, изучаются особенности течения этого процесса при различных состояниях и заболеваниях.

В настоящее время не вызывает большого сомнения сам факт развития системного воспаления неинфекционной природы после множественных травм, больших полостных хирургических операций, в том числе выполненных в условиях искусственного кровообращения, инсультов и т.д. Однако отношение к этому процессу неоднозначное. С одной стороны, системный воспалительный ответ – это реакция организма на воздействие высокой силы (как правило, местное, но с дистантными эффектами) и в таком случае эту реакцию можно считать нормальной, физиологичной. С другой стороны, в ряде случаев СВО рассматривается как патологическая реакция в случаях, когда организм не может локализовать местное повреждение, что приводит к генерализации процесса, повреждении внутренних органов, развитию полиорганной недостаточности.